点击蓝字 关注我们
基于光亲和标记策略鉴定青蒿素在恶性疟原虫红细胞内期中的抗疟靶标

iMeta主页:http://www.imeta.science
研究论文
● 原文链接DOI: https://doi.org/10.1002/imt2.176
● 2024年2月19日,中国中医科学院青蒿素研究中心王继刚、徐承超,南方科技大学戴凌云和河南大学药学院韩光等团队在iMeta在线联合发表了题为 “Photoaffinity probe-based antimalarial target identification of artemisinin in the intraerythrocytic developmental cycle of Plasmodium falciparum” 的研究文章。
● 本研究利用光亲和标记策略鉴定了青蒿素在疟原虫红细胞内期不同阶段的蛋白靶标。结果表明,青蒿素在疟原虫体内可以通过共价和非共价两种结合方式与疟原虫蛋白相互作用,为青蒿素的抗疟作用机制及其蛋白靶点研究提供了新的见解。
● 第一作者:高鹏、王建友、邱崇、张会敏
● 通讯作者:王继刚(jgwang@icmm.ac.cn)、戴凌云(lingyun.dai@outlook.com)、徐承超(ccxu@icmm.ac.cn)、韩光(hang@henu.edu.cn)
● 合作作者:王晨、张迎、孙鹏、陈虹霖、Yin Kwan Wong、陈嘉鋆、张珺哲、唐欢、史巧丽、朱永平、沈胜楠
● 主要单位:中国中医科学院、深圳市人民医院、河南大学、山东省中医药研究院
亮 点
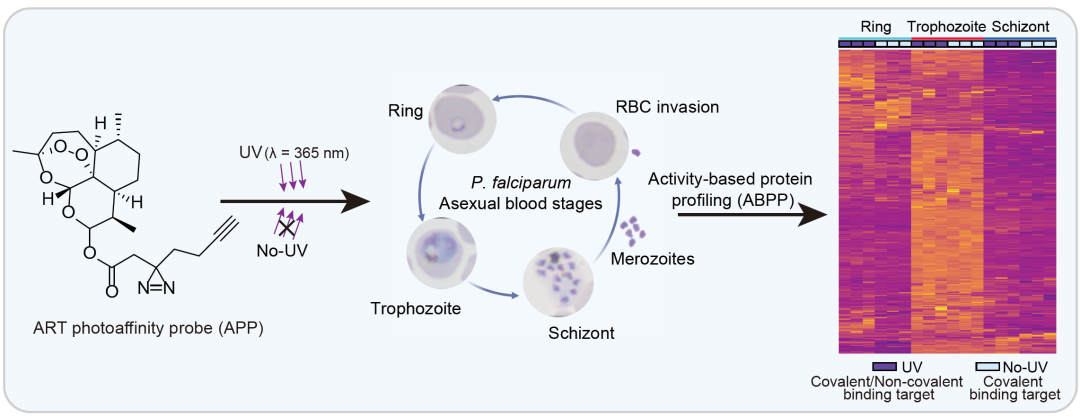
● 利用青蒿素光亲和探针鉴定了青蒿素在恶性疟原虫红细胞内期不同生长阶段的靶标蛋白;
● 青蒿素在疟原虫体内可以通过共价和非共价机制与疟原虫蛋白相互作用;
● 青蒿素可能通过干扰疟原虫的蛋白合成、糖酵解和氧化还原稳态通路发挥其抗疟作用;
● 本研究为探究青蒿素抗疟靶标及其作用机制提供了新的见解。
摘 要
目前,疟疾仍然对全球健康构成严重威胁,青蒿素依然是治疗疟疾的核心药物。然而,由于青蒿素耐药现象的出现和传播,疟疾的防治形势日益严峻。近年来,对青蒿素抗疟作用机制的研究虽然取得了重大进展,但是这些研究主要集中在青蒿素被血红素激活以后的共价结合蛋白靶标上,对青蒿素在疟原虫中的可逆非共价结合靶标的研究较少,并且缺乏有关青蒿素在疟原虫红细胞内期不同生长阶段的靶标信息。本研究利用光亲和标记策略鉴定了青蒿素在疟原虫红细胞内期不同阶段的蛋白靶标。结果表明,青蒿素在疟原虫体内可以通过共价和非共价两种结合方式与疟原虫蛋白相互作用。随后的生物学验证包括靶点验证、表型研究以及非靶向代谢组分析结果表明干扰疟原虫蛋白质合成、糖酵解和氧化还原稳态是青蒿素发挥抗疟活性的关键机制。综上所述,本研究为青蒿素的抗疟作用机制及其蛋白靶点研究提供了新的见解。
视频解读
Bilibili:https://www.bilibili.com/video/BV1Qy421e7sN
Youtube:https://youtu.be/MY0QD0aj9Fs
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
疟疾是一种致命的传染性疾病,2021年在全球造成超过619,000人死亡,大部分是由恶性疟原虫感染所致。恶性疟原虫具有复杂漫长的生命周期,这可以帮助其适应不同的环境变化来逃避防治。恶性疟原虫的生命周期主要可以分为红细胞内期和红细胞外期两个阶段,其中48小时发育周期的致病性红细胞内期(IDC)根据形态特征可以分为环期、滋养体时期和裂殖体时期。当前,大部分药物的作用主要针对疟原虫的红细胞内期,特别是滋养体时期,是大多数抗疟药物的主要作用时期。青蒿素(ART)是为数不多的在疟原虫的整个无性IDC阶段都有效的抗疟药物之一。
通过推广青蒿素和以青蒿素为基础的联合疗法(ACTs),疟疾防治取得了重大进展。尽管ART仍然是推荐的一线抗疟药物,但在东南亚和非洲,人们对ART和ACTs开始产生耐药性的担忧不断加剧。近些年来,多个团队使用多种技术对ART的抗疟机制进行了广泛研究。我们团队主要利用基于活性的蛋白质分析(ABPP)方法来鉴定ART的靶标,并揭示了ART特异的抗疟活性是由于在疟原虫中血红素有效地激活了ART,从而混杂地靶向作用于疟原虫蛋白的机制。虽然我们和其他小组已经鉴定出被血红素激活的ART自由基烷基化的共价结合靶标,但是一些研究表明ART的某些药理作用可能不依赖于ART的共价结合。事实上,我们最近的工作表明ART可以在体外与少数疟原虫蛋白发生非共价相互作用。先前的研究表明,疟原虫在IDC的不同发育阶段(即环期、滋养体期和裂殖体期)对ART表现出不同的敏感性,这表明ART结合的蛋白在不同阶段可能有所不同。此外,血红素介导的ART激活模式也预示着ART靶标蛋白在IDC的不同阶段可能有所不同,因为疟原虫消耗血红蛋白(Hb)和释放血红素的速率在不同阶段是有差异的。然而,之前关于ART靶点的研究似乎忽略了这些方面。因此,有必要全面识别红细胞内期疟原虫不同阶段的ART靶点,以进一步提高我们对ART抗疟机制的认识。
如图1A所示,在本研究中,我们基于ABPP技术利用ART光亲和探针(APP)鉴定了ART在环期、滋养体和裂殖体阶段的相互作用蛋白靶标(包括共价和非共价结合靶标)。APP的抗疟活力与青蒿琥酯(ATS)相当,ATS是ART衍生物,具有更好的水溶性。在APP的结构中引入了双吖丙啶光活性基团和炔烃报告基团,在暴露于365 nm紫外线 (UV) 照射和随后的点击化学反应后能够与靶标发生共价结合,从而有利于同时捕获共价和非共价靶标。然后通过靶点验证、表型研究和非靶向代谢组学分析来进行综合的机制研究。结果表明,ART可以以共价或非共价方式与许多疟原虫蛋白相互作用,这可能共同导致疟原虫的蛋白合成、糖酵解能量供应和氧化还原稳态的破坏,最终导致其显著的抗疟作用。
结 果
恶性疟原虫IDC不同阶段APP的荧光标记
为了研究ART在恶性疟原虫IDC中的抗疟靶点,我们首先确认了APP抗疟活性,然后使用APP在有或没有紫外线照射的情况下对疟原虫进行原位荧光标记。结果表明,在两种条件下APP对疟原虫蛋白的标记均具有剂量依赖性,并且紫外线照射对标记效率影响很小。竞争实验进一步证明了探针的特异性。然而,考虑到ART独特的血红素介导的激活模式和疟原虫生命周期的复杂性(图1B),我们接下来验证了APP和疟原虫蛋白在IDC的环期、滋养体和裂殖体阶段之间的相互作用(图1C)。结果如图1D所示,APP可以与三个不同阶段疟原虫产生相互作用,表明ART可以在整个IDC时期都可以被血红素激活。滋养体阶段的荧光标记强度明显强于环期和裂殖体阶段(图1D),表明ART在滋养体阶段激活程度更高,可以结合更多的疟原虫蛋白。在活细胞成像实验中,我们使用共焦显微镜观察了APP在环期、滋养体和裂殖体阶段的分布(图1E)。结果表明,APP在滋养体阶段疟原虫中结合了更多的蛋白,与凝胶中的荧光标记结果一致。
此外,我们还发现,环期和裂殖体时期疟原虫在紫外线照射后荧光标记强度增强(图1D),这表明除了自由基反应的不可逆共价结合模式外,ART还可以与某些蛋白质以非共价模式相互作用,或者同时以这两种方式与相同的靶标蛋白产生结合。然而,紫外线照射后滋养体阶段的荧光强度没有显著差异(图1D)。这可能是因为疟原虫在滋养体阶段消耗更多的血红蛋白(Hb),产生浓度更高的血红素,ART被迅速激活,从而在光交联反应发生之前将大部分靶标蛋白烷基化。另一种可能是由共价相互作用引起的蛋白质构象变化阻止了非共价结合。
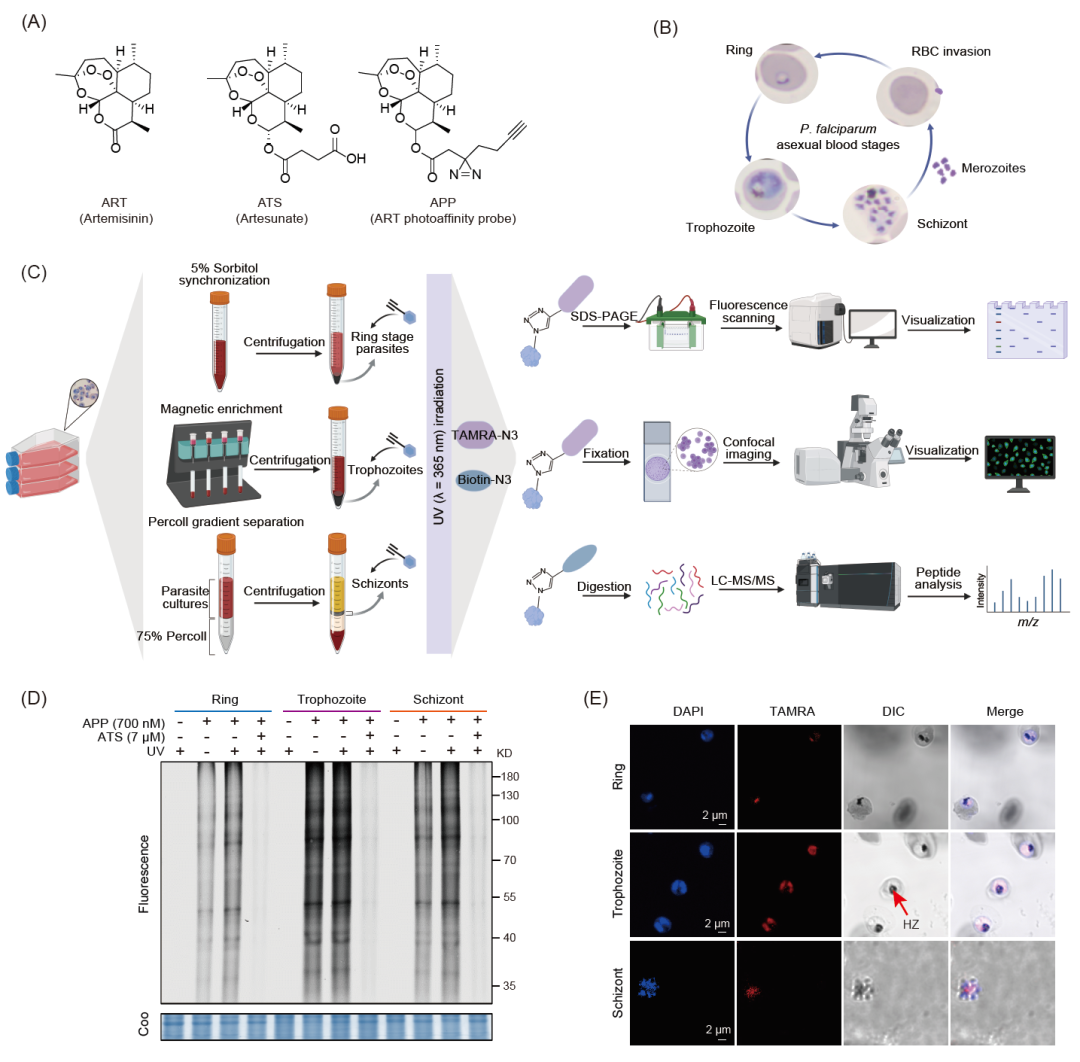
图 1. 恶性疟原虫环体、滋养体和分裂体阶段光亲和APP的原位荧光标记
(A)青蒿素(ART)、青蒿琥酯(ATS,一种水溶性更好的ART衍生物)和青蒿素光亲和活性探针(APP)的化学结构。(B)恶性疟原虫红细胞内期生长发育(IDC)示意图。裂殖体侵入红细胞,经历环期、滋养体和裂殖体阶段完成整个IDC。(C)用于标记和鉴定体内ART靶标蛋白的光亲和探针APP的基于活性的蛋白质组分析(ABPP)的一般工作流程。(D)恶性疟原虫不同阶段APP的原位标记及与过量ATS的竞争。(E)共聚焦成像显示恶性疟原虫不同阶段APP (700 nmol/L)在紫外线照射下的分布(比例尺= 2 µm)。APP,青蒿素光亲和探针; P. falciparum, 恶性疟原虫; HZ, 疟原虫色素; Coo,考马斯亮蓝; nRBC,正常红细胞; UV, 紫外线; LC-MS/MS,液相色谱-串联质谱; KD, kilo-Dalton; DAPI, 4’,6-Diamidino-2-phenylindole; TAMRA, carboxytetramethylrhodamine; DIC, 恶性疟原虫红细胞内期生长发育周期。
IDC不同阶段APP靶标蛋白的鉴定
受上述荧光标记结果的启发,我们随后尝试在紫外线照射或不照的情况下,使用pull-down鉴定APP在恶性疟原虫的环期、滋养体和裂殖体阶段的靶标蛋白。如图1C所示,将不同阶段的疟原虫与APP或二甲基亚砜(DMSO)对照组一起孵育,随后进行或不进行紫外线照射。然后,进行点击化学反应,将标记的蛋白质与生物素标签结合,用链霉亲和素珠子富集目标蛋白,用胰蛋白酶消化,最后通过高分辨率质谱(MS)进行鉴定。如图2A所示,共有451个潜在的靶标蛋白被鉴定到,其中247、396和353个蛋白分别在恶性疟原虫的环期、滋养体和裂殖体时期被鉴定。在这451个靶标中,177个靶标在所有三个阶段都被鉴定出来。滋养体阶段的靶标数量最多,这也解释了滋养体阶段是IDC中对ART最敏感的阶段。对结合蛋白丰度比的分析清楚地表明,环期和裂殖体阶段,紫外线照射下一些目标蛋白的信号相对高于非紫外线照射下的信号(图2B)。然而,在滋养体阶段,紫外线照射对蛋白相对丰度的影响要小得多(图2B)。因此,结果表明,除了血红素激活介导的与蛋白的共价结合外,ART与疟原虫蛋白的非共价结合也会发生。
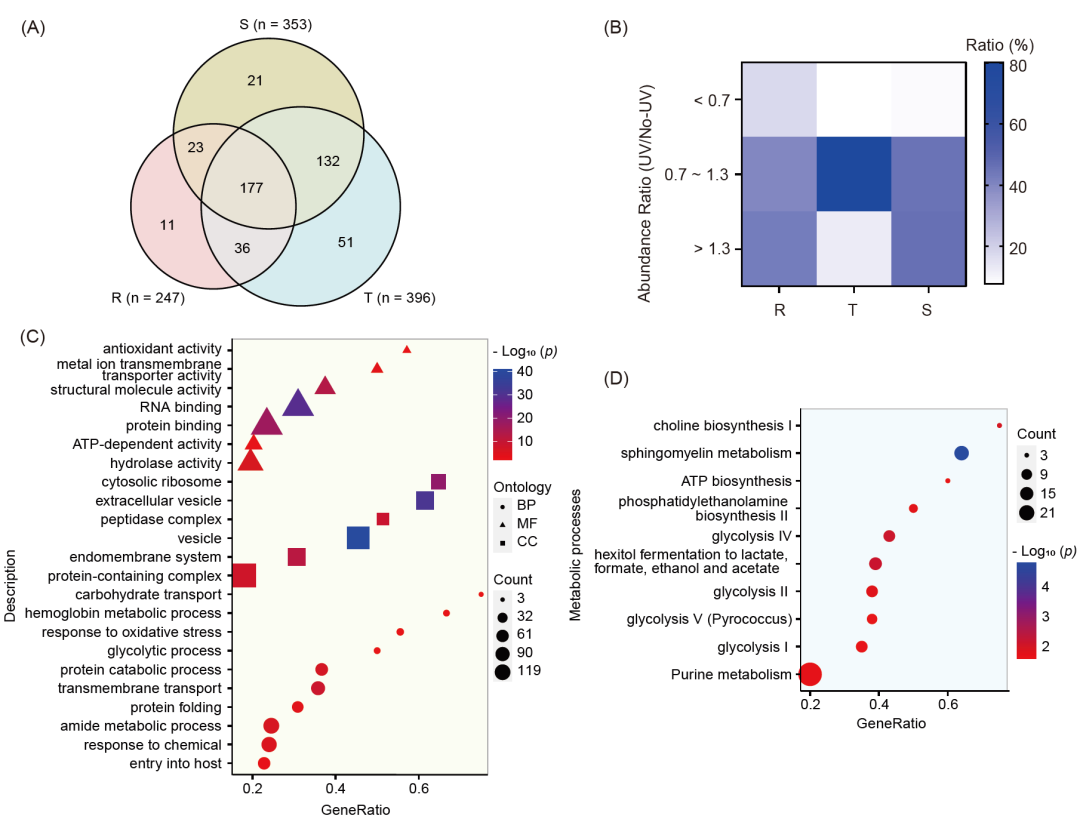
图2. 基于ABPP技术利用APP鉴定ART的靶标蛋白和富集分析
(A) APP在恶性疟原虫不同阶段鉴定的靶标蛋白的维恩图。(B) APP原位鉴定不同时期恶性疟原虫靶标蛋白的丰度比。(C) 451个靶点的代谢途径富集分析。(D)所有451个靶点的基因本体(GO)富集分析。包括生物过程(BP)、分子功能(MF)和细胞组成(CC)三大类。R, 环期; T, 滋养体; S, 裂殖体。UV,紫外线照射; No-UV,无紫外线照射。
靶标蛋白质的生物信息学分析
接下来,我们对这451个靶标进行了基因本体(Gene Ontology,GO)分析,结果表明这些靶标具有多种生理功能(分子功能(Molecular Function,MF)分析),并参与了疟原虫体内不同亚细胞定位(细胞组分(Cellular Component,CC)分析)的多种生物过程(生物过程(Biological Process,BP)分析)(图 2C)。此外,我们还对这些靶标可能参与的代谢过程进行了富集分析。如图2D所示,结果表明这些靶标主要参与了几种代谢相关的通路,包括糖酵解以及胆碱、磷脂和嘌呤的生物合成和代谢。蛋白质-蛋白质相互作用(PPI)分析表明,鉴定出的靶标主要参与转录、蛋白质折叠和分解代谢等多个过程(图 S4)。
我们还对三个阶段的靶标蛋白进行了GO富集分析。分析结果显示,环期和滋养体时期的靶标蛋白主要参与疟原虫蛋白质分解、水解等过程,而裂殖体时期的靶标蛋白主要参与肽代谢、酰胺生物合成、翻译等过程(图3A-C)。此外,我们还发现四个生理过程,包括翻译、蛋白水解、肽代谢和生物合成过程,在三个不同阶段都有富集(图3D)。然而,不同阶段的靶向程度各不相同,在环期和滋养体阶段的疟原虫中观察到的靶向程度最高。这表明ART对特定生理过程的影响在不同阶段会有波动,这可能是由于ART在环期、滋养体和裂殖体阶段的激活水平不同,因此靶标蛋白的共价和非共价结合也不同。
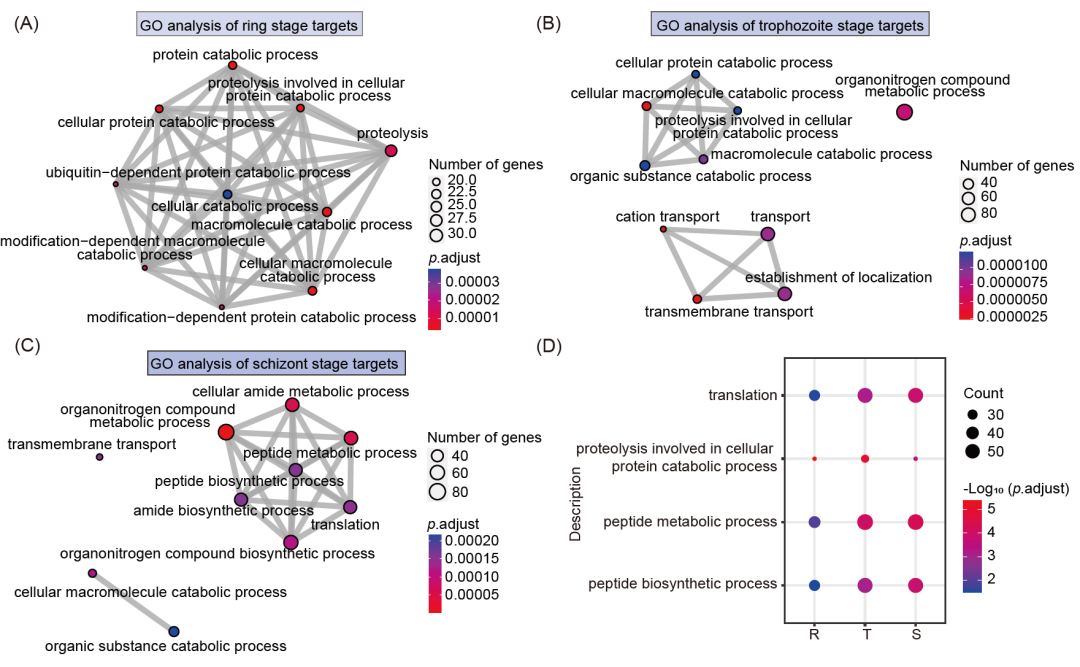
图3. 鉴定的APP靶标蛋白生物信息学分析
(A)环期靶标蛋白的BP网络分析。(B)滋养体时期靶标蛋白的BP网络分析。(C)裂殖体时期靶标蛋白的BP网络分析。(D)共同的生物学过程在IDC的三个不同阶段得到的富集。
青蒿素干扰恶性疟原虫的新生蛋白合成
靶标蛋白分析表明,干扰翻译和新生蛋白合成相关过程可能是ART产生抗疟作用的关键途径。在恶性疟原虫的IDC过程中,疟原虫会消化高达95%的宿主红细胞Hb,并释放出大量氨基酸用于蛋白质合成,同时翻译和增殖也十分活跃。因此,我们接下来验证了ATS是否会抑制疟原虫的蛋白合成。L-azidohomoalanine(AHA)是一种非放射性 L-蛋氨酸类似物,可在蛋白合成过程中掺入蛋白结构中,我们用它来监测ATS对疟原虫新生蛋白从头合成的影响。AHA的叠氮部分可以与生物素-炔探针发生点击化学反应,然后通过亲和珠富集,胰酶消化后通过高分辨率MS鉴定。如图4A所示,与对照组相比,ATS处理后 1421种蛋白的合成受到明显抑制。这些蛋白主要参与有机物质分解、细胞内转运和其他关键生理过程(图4B)。
恶性疟原虫延伸因子1-α(PfEGF1-α,PF3D7_1357000)被鉴定为ATS在IDC所有三个阶段共有的潜在靶标。随后,我们表达并纯化了重组PfEGF1-α蛋白,并验证了它与APP的相互作用。如图4C和4D所示,在紫外照射下,APP对PfEGF1-α的标记呈现剂量相关性,过量的ATS可与之产生竞争性结合。在原位pull-down免疫印迹试验中也观察到类似的竞争性结合现象(图4E)。药物亲和反应靶点稳定性(DARTS)分析表明,与ATS孵育后,PfEGF1-α对蛋白水解的抵抗力增强(图 4F),表明ATS与PfEGF1-α有特异性结合。免疫荧光结果也显示APP与PfEGF1-α在原位共定位(图4G和4H)。此外,我们还注意到,在与半胱氨酸(Cys)特异性阻断剂碘乙酰胺(IAA)预孵育后,APP的荧光标记强度有不同程度的下降(图4D),这表明Cys可能是ATS与PfEGF1-α结合的氨基酸位点之一。最后,我们研究了不同条件下PfEGF1-α与ATS的结合情况。如图4I所示,仅在紫外照射下,PfEGF1-α的荧光标记强度与仅在血红素和抗坏血酸钠激活下的荧光标记强度相当,这表明ATS能以共价和非共价两种模式与PfEGF1-α结合。
我们还研究了恶性疟原虫整个IDC蛋白质从头合成速率的变化以及ATS对这一过程的影响。结果表明疟原虫蛋白质的合成速率是波动的,其中滋养体阶段的合成速率最高,环期最低,ATS处理后减弱了整个IDC阶段的蛋白质合成过程。我们还分析了环期、滋养体和裂殖体阶段蛋白水解的变化。滋养体阶段和裂殖体阶段的蛋白酶活性高于环期,这表明蛋白水解程度在环期之后有所提高。结果还显示,ATS处理在不同程度上抑制了蛋白酶活性。此外,许多核糖体蛋白也被鉴定为 ART的潜在靶标,进一步证明蛋白质合成可能受到影响。总之,这些结果表明,ART可能会在整个IDC中与包括PfEGF1-α在内的相关疟原虫蛋白结合,从而抑制蛋白质合成,进而产生抗疟效果。
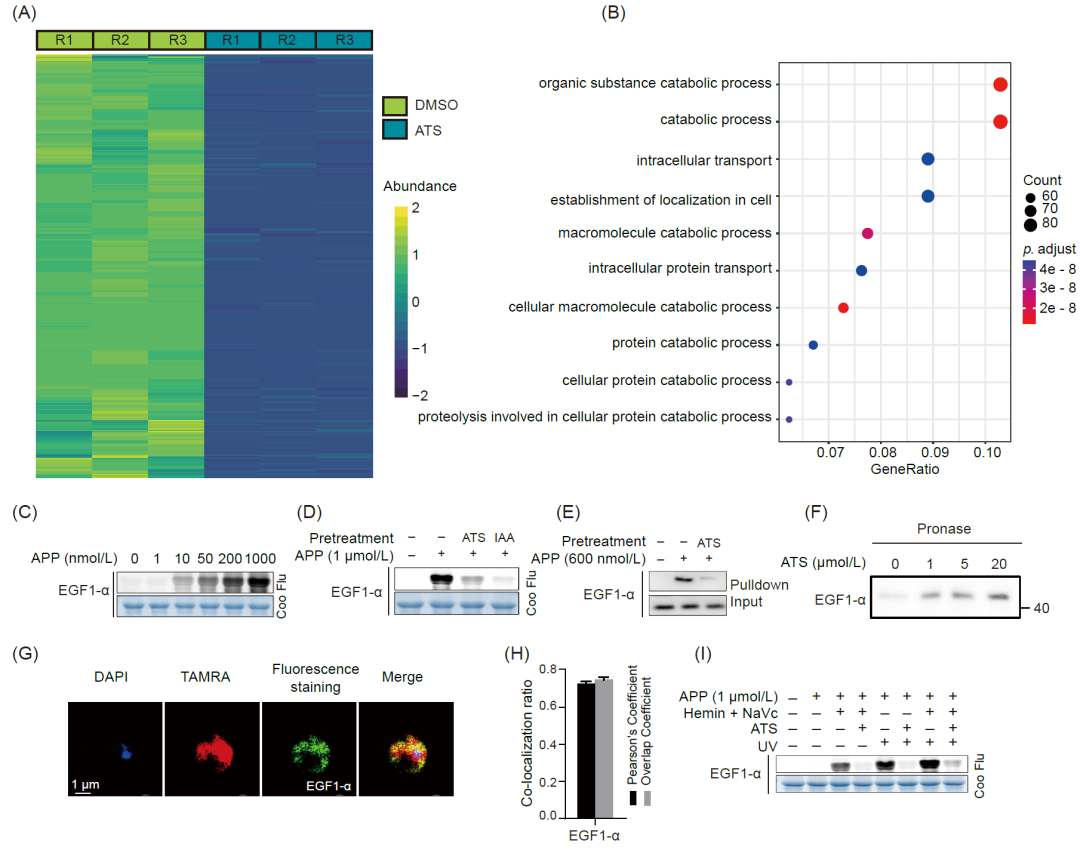
图 4. 青蒿素干扰疟原虫的蛋白质合成
(A)与对照组(DMSO)相比,ATS处理后合成降低的疟原虫蛋白热图。(B)合成降低的蛋白的GO-BP富集分析。(C)APP对重组PfEGF1-α (PF3D7_1357000)进行剂量依赖性荧光标记。(D)过量ATS (10 μM)和IAA (10 μM)预孵育与重组蛋白的APP标记竞争。(E) APP原位结合PfEGF1-α的Pull-down免疫印迹验证。(F)使用药物亲和反应的靶点稳定性(DARTS)方法验证ATS对靶蛋白的作用。(G)紫外照射下APP与PfEGF1-α蛋白共定位的免疫荧光染色代表图。(H) (G)共定位的定量分析。(I)不同条件下重组PfEGF1-α用APP荧光标记。EGF1-α,延伸因子1-α; IAA, 碘乙酰胺。
青蒿素干扰恶性疟原虫的糖酵解过程
正如之前的靶标蛋白生信分析结果所示,与糖酵解和能量代谢有关的主要代谢途径被显著富集,这时因为恶性疟原虫在IDC期间需要消耗大量葡萄糖,并严重依赖糖酵解为快速生长和增殖提供能量。这也导致丙酮酸和乳酸的产生增加。糖酵解途径已被确定为ART产生抗疟活性的必要途径。因此,我们继续研究ART对疟原虫糖酵解的影响。首先,我们检测了疟原虫体内的丙酮酸水平以及疟原虫和培养基中的乳酸水平。结果表明,随着药物浓度的增加,丙酮酸和乳酸的水平显著下降(图5A-C)。ATP水平也呈同样的下降趋势(图5D)。然后,我们通过测量细胞外酸化率(ECAR),进一步评估了ATS对红细胞内寄生虫糖酵解活性的影响。如图5E和F所示,ATS以剂量依赖的方式显著削弱了疟原虫的糖酵解过程和糖酵解能力,这与乳酸水平的降低是一致的。
考虑到糖酵解是最基本的代谢途径之一,与之相关的一些高活性、高表达的酶被认为是抗疟药物开发的有潜力的靶标。在我们之前的研究中,恶性疟原虫L-乳酸脱氢酶(PfLDH,PF3D7_1324900)、磷酸甘油醛异构酶(PfTIM,PF3D7_1439900)和甘油醛-3-磷酸脱氢酶(PfGAPDH,PF3D7_1462800)被确定为青蒿素的潜在靶标,本研究也对它们进行了鉴定。我们首先利用DARTS验证了它们与ATS的相互作用,并测定了ATS对它们酶活性的抑制作用,结果表明ATS在低浓度下即可产生抑制作用。此外,我们还尝试利用高分辨率质谱鉴定 ATS与重组PfGAPDH的可能共价结合位点。结果表明,谷氨酸残基59(Glu 59)可能是一个结合位点,分子对接模拟也证实了这一点。上述结果表明,ATS可能与糖酵解酶(包括 PfLDH、PfTIM 和 PfGAPDH)结合,抑制它们的活性,干扰疟原虫的糖酵解过程,进而在IDC期间发挥抗疟作用。
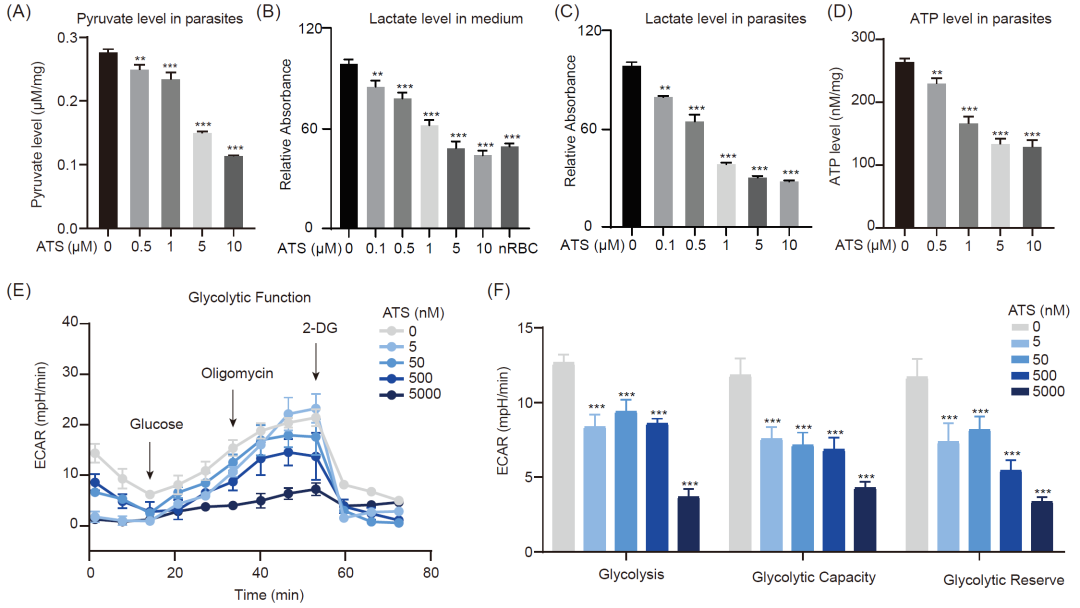
图 5. 青蒿素干扰疟原虫的糖酵解过程
(A) ATS降低疟原虫体内丙酮酸水平。(B) ATS以剂量依赖的方式降低培养基中乳酸的水平。(C) ATS以剂量依赖的方式降低疟原虫体内乳酸水平。(D) ATS降低疟原虫体内ATP水平。(E) Seahorse海马代谢仪测定ATS对糖酵解活性的抑制作用。(F) ATS以剂量依赖性方式抑制恶性疟原虫的糖酵解活性。所有数据均取三个独立实验的平均值,所有数值均以均数±标准差(SEM)表示(*p < 0.05,**p < 0.01,***p < 0.01)。ECAR, 细胞外酸化速率; 2-DG, 2-脱氧-D-葡萄糖; ATP, 5'-三磷酸腺苷。
青蒿素干扰恶性疟原虫的氧化还原稳态
在IDC期间,疟原虫消耗大量葡萄糖来提供能量,并降解大量Hb来促进生长和增殖。这一代谢过程会产生大量氧化副产物,包括超氧阴离子自由基(O2˙-)和过氧化氢(H2O2)。由此产生的氧化损伤使疟原虫处于高度氧化应激状态。如图2D所示,在对所有靶标的GO分析中,抗氧化活性和氧化应激途径都被显著富集。疟原虫的抗氧化防御系统主要包括谷胱甘肽(GSH)和各种硫代氧化酶依赖蛋白。谷胱甘肽是疟原虫体内含量最高的低分子量氧化还原活性硫醇,可中和H2O2等氧化副产物,有效维持细胞内的氧化还原稳态。此外,GSH水平的波动也可能影响ART的药物敏感性。在这里,我们同时检测了疟原虫体内的H2O2和GSH水平。如图6A、B所示,在ATS处理后,H2O2显著增加,而GSH则呈现相反的趋势。这些结果表明,ATS可能会通过破坏疟原虫的氧化还原平衡而对其造成氧化损伤,ATS处理后疟原虫体内丙二醛和过氧化物酶活性的变化也证明了这一点。此外,我们还发现活化的ATS可在体外与GSH的硫醇基发生共价结合。
蛋白质S-谷胱甘肽化反应会影响修饰蛋白质的功能(图6C)。一个重要的例子是恶性疟原虫鸟氨酸氨基转移酶(PfOAT,PF3D7_0608800),它通过调节鸟氨酸水平直接影响多胺的合成,并参与葡萄糖代谢和细胞增殖的调节。PfOAT的活性位点半胱氨酸残基154和163可通过 S-谷胱甘肽化修饰形成二硫键,导致底物结合障碍,从而影响催化活性。在这项研究中,PfOAT在IDC所有三个阶段中都被鉴定为潜在靶点,而且在其他研究中也被确定过。我们首先在体外验证了APP与PfOAT的特异性结合(图6D-I)以及两种结合模式的存在(图6J)。酶活性实验表明,ATS可显著抑制血红素激活和非激活状态下的酶活性(图 6K、L),并降低疟原虫体内鸟氨酸的水平(图6M)。这些结果都清楚地表明,共价和非共价结合模式可能共同参与了ATS与PfOAT的相互作用。
此外,我们还验证了APP与两种参与维持疟原虫体内氧化还原平衡的重要过氧化蛋白—恶性疟原虫硫过氧化物酶-1(PfTrx-Px1,PF3D7_1438900)和1-Cys过氧化还原酶(Pf1-CysPxn,PF3D7_0802200)的相互作用,这两种蛋白也是本研究中发现的潜在结合靶标之一。结果表明,ATS也可能与这些蛋白质发生共价或非共价结合,从而影响疟原虫的氧化还原平衡。
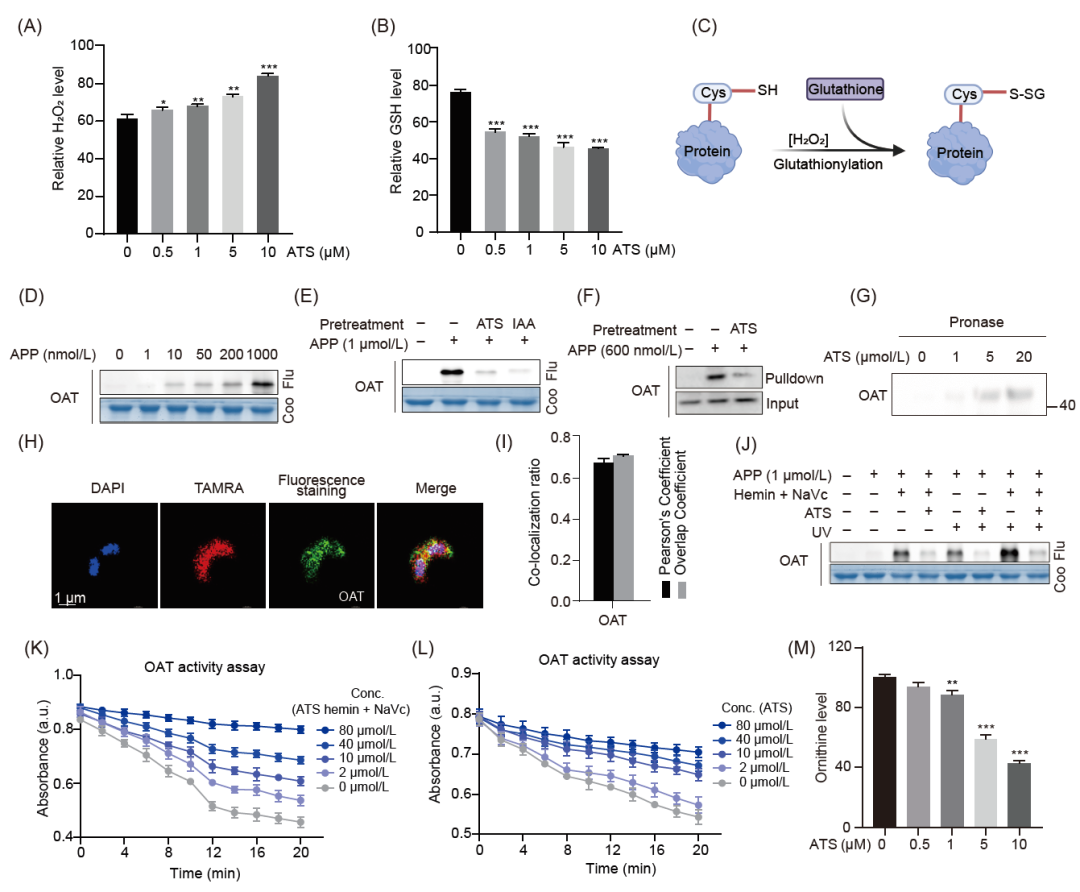
图 6. H2O2、GSH的测定及ATS靶向PfOAT (PF3D7_0608800)的验证
(A) ATS增加恶性疟原虫H2O2水平。(B) ATS降低恶性疟原虫谷胱甘肽水平。(C)蛋白质S-谷胱甘肽化示意图。(D)APP对重组PfOAT (PF3D7_1357000)进行剂量依赖性荧光标记。(E)过量ATS (10 μM)和IAA (10 μM)预孵育与重组蛋白的APP标记竞争。(F) APP与PfOAT原位结合的Pull-down免疫印迹验证。(G) 使用药物亲和反应的靶点稳定性(DARTS)方法验证ATS对靶蛋白的作用。(H)紫外照射下APP与PfOAT蛋白共定位的免疫荧光染色代表图。(I) (G)共定位的定量分析。(J)不同条件下重组PfOAT的APP荧光标记。(K) ATS在体外以剂量依赖的方式抑制重组PfOAT的酶活性,并伴有血红素激活。(L) ATS体外抑制重组PfOAT酶活性呈剂量依赖性,无血红素激活。(M) ATS以剂量依赖的方式降低鸟氨酸水平。所有数据均基于至少3个独立的生物重复,以均数±SEM表示(*p < 0.05,**p < 0.01,***p < 0.01)。Flu,荧光; Conc, 浓度; a.u., 相对单位; GSH, 谷胱甘肽; OAT, 鸟氨酸转氨酶。
青蒿素干扰恶性疟原虫的代谢过程
当疟原虫处于巨大的外来压力下并且不能代偿时,会影响包括代谢途径在内的多个系统的级联反应。鉴于ATS对疟原虫的糖酵解途径有明显的抑制作用,以及ATS对疟原虫的各种生理过程的干扰,我们接下来使用非靶向代谢组学分析来研究ATS对疟原虫代谢组的影响(图7A)。
首先,我们进行了偏最小二乘判别分析(PLS-DA),评估对照组和ATS处理的疟原虫之间的代谢差异。结果显示,两组疟原虫之间存在明显差异,表明ATS对疟原虫的新陈代谢具有显著影响。非靶向代谢组学图谱共鉴定出818个代谢物用于进一步分析。利用变量重要度(VIP) > 1、P值 < 0.05 和倍数变化(FC) > 1.2 来筛选显著改变的代谢物,一共发现42个差异表达的代谢物,其中,30个代谢物下调,12个上调(图7B)。其中氧化型谷胱甘肽水平显著增加(图7C),与图6B中的测量结果一致。此外,还观察到包括LPS和PC在内的脂质代谢相关产物显著减少(图7D),这与疟原虫在红细胞内期需要大量脂质物质来构建细胞膜用以新生疟原虫生长的情况一致。最后,我们将APP鉴定得到的靶标蛋白映射到富集的代谢过程中,并确定了参与各代谢过程的一些潜在的最重要靶标蛋白(图 7E)。由于疟原虫的单倍体性质以及许多疟原虫蛋白是由单拷贝基因编码的,可以认为大多数定位的蛋白是与ATS影响疟原虫代谢相关的关键靶点。
综上所述,我们的非靶向代谢组学分析表明ATS可以广泛地干扰红细胞内期疟原虫的多种代谢途径,这些途径可能直接或间接地汇聚到ART的综合抗疟作用上。
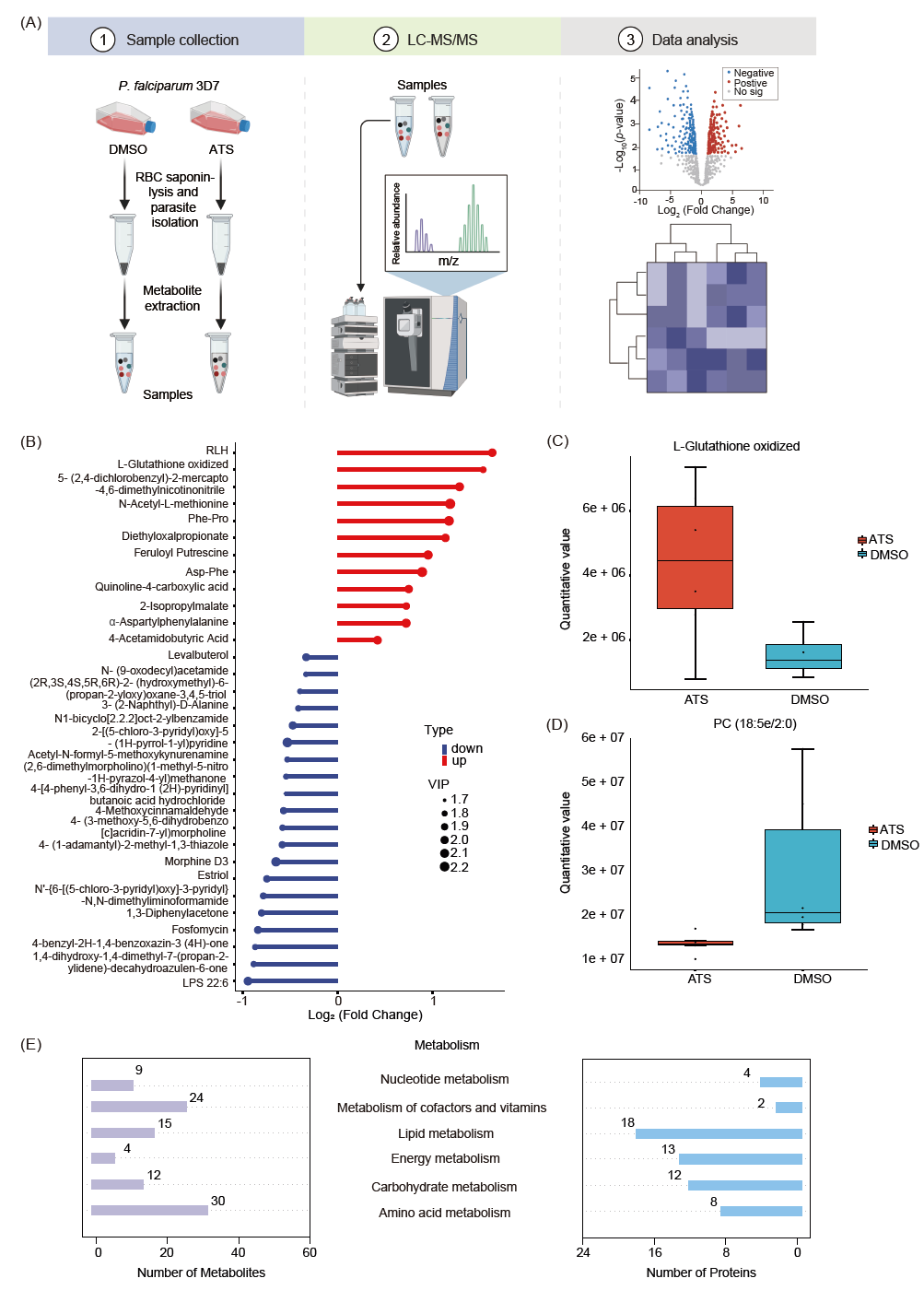
图7. 与对照相比,ATS处理恶性疟原虫的非靶向代谢组学分析
(A)非靶向代谢组学研究的工作流程示意图,包括样品收集、LC-MS/MS鉴定和数据分析。(B)杆图显示有显著差异的代谢物。(C)与对照组(DMSO)相比,ATS治疗后氧化型L-谷胱甘肽水平。(D)与对照组(DMSO)相比,ATS处理后氧化PC (18:5e/2:0)水平。(E)参与富集代谢通路的代谢物和相关蛋白。数据以平均值±均值标准差表示。
方 法
恶性疟原虫培养
恶性疟原虫3D7虫株的连续培养是在先前的方法基础上稍作修改的。疟原虫在补充有2%健康人红细胞的完全培养基中培养,并在摇床上轻微摇动。每天制备吉姆萨染色的薄血涂片评估疟原虫感染率。关于环期、滋养体和裂殖体高度同步化的疟原虫富集,是首先使用5%山梨醇将疟原虫紧密同步两次获得环期疟原虫,然后通过磁力分离(MACS CS柱,Miltenyi Biotech)富集获得滋养体疟原虫,最后裂殖体时期疟原虫通过70%Percoll(v/v)缓冲液离心富集得到。
恶性疟原虫原位荧光标记
原位荧光标记是根据我们之前描述的方法进行的,并稍作修改。疟原虫在6孔板中以5%的感染率培养。与700nm APP或等量溶剂(DMSO,终浓度小于0.1%)孵育4小时后,在冰面上暴露或不暴露于365 nm紫外线照射10分钟。对于竞争实验,疟原虫用过量的ATS预处理1小时,然后用APP处理4小时。疟原虫从红细胞中裂解释放后,超声提取可溶性疟原虫蛋白,并取等量的蛋白溶液(20µg)进行点击化学反应(CuAAC)用四甲基-6-羧基罗丹明叠氮 (TAMRA-azide)(50μM)进行荧光标记。点击化学反应结束后,用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳法(SDS-PAGE)分离蛋白质进行荧光成像。
ART结合靶标蛋白鉴定
Pull-down实验按照之前的文献进行。用700 nM APP或等体积的DMSO处理不同红细胞内期的高度同步化疟原虫4小时。然后在冰上或不在冰上用365 nm紫外线照射样品10分钟。照射后,按上述方法提取可溶性疟原虫蛋白并进行定量。紧接着进行CuAAC反应,用生物素叠氮化物将生物素与APP连接。过夜沉淀蛋白和重新溶解后,用链霉亲和素珠子(美国 Thermo 公司)富集蛋白质。为了进行蛋白鉴定,分别用二硫苏糖醇(DTT)和碘乙酰胺(IAA)对富集的蛋白质进行还原和烷基化,并用胰蛋白酶(Promega,美国)在37°C下消化过夜。然后离心收集上清液,脱盐并干燥,用于随后的液相色谱-串联质(LC-MS/MS)分析。
关于目标蛋白的免疫印迹验证,用SDS-PAGE对富集的蛋白进行分离。然后将蛋白转移到聚偏二氟乙烯(PVDF)膜上,与链霉亲和素-HRP或适当的一抗和二抗孵育,并使用电化学发光(ECL)显像。
LC-MS/MS 检测和蛋白质鉴定
靶标鉴定的LC-MS/MS分析如前所述。数据采集使用XCalibur(版本4.2.4)以数据依赖采集模式进行。所有光谱都被检测到,MS光谱在300-1500 m/z之间,分辨率为60,000。利用SEQUEST HT与恶性疟原虫3D7蛋白数据库(PlasmoDB-60,https://plasmodb.org/plasmo/app/downloads/release60/P.falciparum3D7/fasta/data/)进行比对。如前所述,设置静态和动态修改。
APP的细胞内成像分析
细胞成像实验按照先前研究中的描述进行[43]。用APP或DMSO处理疟原虫30分钟,固定后滴加在预先包被有多聚赖氨酸的盖玻片上打孔。然后与TAMRA-azide进行点击化学反应。随后,将盖玻片转移到含有4’,6-二氨基苯基吲哚(DAPI)涂层的载玻片上,使用TCS SP8SR共聚焦显微镜进行成像。
ATS对恶性疟原虫新生蛋白质合成的影响
L-叠氮基高丙氨酸(AHA)用于鉴定ATS抑制的新生蛋白。在预先30分钟消耗掉细胞内储存蛋氨酸后,用ATS(700 nM)或溶剂对照处理非同步化疟原虫,在含AHA的无蛋氨酸培养基中培养6小时。随后,提取疟原虫可溶性蛋白,用CuAAC反应将生物素与AHA标记蛋白偶联,并通过Pull-down实验进行富集和质谱鉴定。
为了进一步验证ATS对疟原虫新生蛋白的抑制作用,实验方法如前所述进行,并作了微小的修改。连续两次使用5% D-山梨醇对环期疟原虫进行高度同步化。含疟原虫的感染红细胞用L-蛋氨酸的培养基洗3次。然后放在培养箱中孵育30分钟,以耗尽细胞内蛋氨酸储备。随后培养基用含AHA的无L-无蛋氨酸培养替代,并与ATS(100 nM)或等体积的溶剂对照反应。每隔5小时收集一次疟原虫样本。最后提取疟原虫可溶性蛋白进行CuAAC反应,十二烷基硫酸钠-PAGE分离后进行荧光成像。
药物亲和反应的靶点稳定性分析
药物亲和反应的靶点稳定性(DARTS)法用于检测ATS与靶标蛋白的结合。用一系列ATS处理重组蛋白(5 μM)并在室温下孵育2小时。然后加入1 μg/mL的链霉蛋白酶,室温孵育10分钟。随后加入上样缓冲液,在95℃下终止反应,进行SDS-PAGE电泳和免疫印迹。
蛋白水解酶活性测定
使用荧光蛋白酶检测试剂盒(G-Biosciences,C006028)测量处于环期、滋养体和裂殖体时期的疟原虫的蛋白酶活性。将处于不同周期的高度同步化疟原虫与ATS(100 nM)孵育4小时。然后收集红细胞,将疟原虫从受感染的红细胞中释放出来,并在冰上超声裂解提取可溶性疟原虫蛋白。在BCA蛋白浓度定量后,取等量蛋白质与蛋白酶底物在37℃下孵育。孵育后,加入沉淀剂再孵育10分钟,然后在12,000×g下离心5分钟。将上清液转移到新的EP管中,并与检测缓冲液混合。在570nm波长处测量吸光度(OD)。
乳酸水平检测
用不同浓度的ATS处理疟原虫4小时。反应结束后收集培养基上清液和感染的红细胞。然后根据乳酸含量检测试剂盒(Solarbio, 中国,BC2235)的指导测定疟原虫乳酸水平。
丙酮酸水平检测
用丙酮酸(PA)含量检测试剂盒(Solarbio, 中国,BC2205)测定疟原虫的丙酮酸水平。用一系列浓度的ATS处理疟原虫4小时,然后用丙酮酸提取缓冲液超声裂解,静置30分钟。在室温下8000g离心10分钟后,收集上清液,根据试剂盒的指导检测丙酮酸。在520 nm波长处检测吸光度(OD)。
细胞能量代谢分析
在Agilent Seahorse XFe96分析仪上使用糖酵解压力测试试剂盒测量恶性疟原虫的细胞外酸化率(ECAR)。所有检测均按先前描述并根据制造商的说明稍作修改后进行。探针板预先在37℃下水化过夜。将恶性疟原虫与浓度不断增加(0-20 µM)的ATS培养,并用0.01% 皂素裂解释放。然后将疟原虫重悬于含有1 mM谷氨酰胺的 Seahorse XF RPMI 1640 培养基(安捷伦科技公司)中,并固定到预先涂有0.01%(w/w)多聚赖氨酸的Seahorse 细胞板中。随后,在相应的加药孔中依次加入相关试剂(10 mM 葡萄糖、1 µM oligomycin和 50 mM 2-DG,并将细胞板和探针板放入海马 XFe96分析仪进行测量。
ATS结合位点的鉴定
如前所述,10 μM重组蛋白与100 μM ATS、20 μM hemin和 200 μM NaVc共孵育4小时。然后用DTT和IAA对样品进行还原和烷基化,并在37°C下消化过夜。最后在离心真空蒸发器中对肽段进行脱盐和旋干,按上述方法进行LC-MS/MS分析。
分子对接模拟
分子对接模拟使用MOE(20190102)进行。ATS的化学结构从PubChem(PubChem ID:6917864)下载。PfGAPDH(PDB:1T24)的蛋白晶体结构从PDB数据库下载,使用QuickPrep模块制备。
过氧化氢和谷胱甘肽含量检测
过氧化氢检测试剂盒(Beyotime, Shanghai, China)和总谷胱甘肽检测试剂盒(Beyotime, Shanghai, China)用于检测疟原虫体内的H2O2和GSH水平。将疟原虫接种到6孔板中,与不同浓度的ATS(0、0.5、1、5 和 10 μM)在37°C培养箱中反应4小时。然后用0.05%的皂素裂解释放疟原虫,并按照试剂盒说明书检测H2O2和GSH的水平。
鸟氨酸氨基转移酶(OAT)活性测定
OAT活性测定方法如前所述。将纯化的PfOAT蛋白(5 μg)与不同浓度(0-80 μM)的ART培养1小时,然后转移到96孔板中,按照试剂盒(Solarbio,中国,BC4405)生产商的说明加入底物NADH和反应缓冲液,在340nm波长处连续检测吸光度。
重组蛋白的表达和纯化
重组蛋白的编码序列下载来自PlasmoDB 数据库(http://PlasmoDB.org)。这些序列由Sangon Biotech公司(中国上海)合成并克隆到pET-28a (+)载体中。重组质粒在大肠杆菌BL21(DE3)中表达,并使用IPTG诱导。然后,在1200 bar的压力下提取含有重组蛋白的细胞裂解液,将其装入Ni-NTA色谱柱,洗脱后获得重组蛋白。
重组蛋白的体外荧光标记
重组靶标蛋白的验证方法如前所述。在剂量依赖性试验中,等量的重组蛋白(2 μg)与浓度不断增加的APP处理4小时。在竞争试验中,重组蛋白先用过量的ATS IAA预处理2小时,再与APP反应2 小时。然后用TAMRA-azide进行 CuAAC。其他步骤与上述步骤相同。
非靶向代谢组样品制备
按上述方法培养恶性疟原虫3D7虫株(未同步化),在2%红细胞压积条件下稀释将疟原虫感染率稀释至10%,和ATS(700 nM)和等体积的溶剂对照(0.1% DMSO)反应6小时。然后收集疟原虫并从红细胞中释放出来,快速将疟原虫放入液氮中速冻,并保存在-80°C,以便在2天内进行进一步的代谢物分析。
UHPLC-MS/MS非靶向代谢组分析和处理
疟原虫的代谢物提取(每个样本2×108个细胞)如前所述。样品被注入Hypersil Gold色谱柱。洗脱液为0.1%甲酸(FA) (A) 和甲醇(B) (正极性模式)和5 mM醋酸铵(pH 9.0) (A)和甲醇(B)(负极性模式)。所有的实验都被独立地重复至少三次。用Compound Discoverer 3.1(ThermoFisher)处理UHPLC-MS/MS产生的原始数据。将峰值强度归一化为总光谱强度,并与mzCloud(https://www.mzcloud.org/)、mzVault和Mass List数据库)进行匹配。以p值 < 0.05、FC > 1.2或FC < 0.833、VIP > 1为筛选差异代谢物的显著阈值。
统计分析
使用 GraphPad Prism 8.3 进行统计分析。所有数据均基于至少三个独立的生物重复,并以平均值 ± 平均值标准误差(SEM)表示。统计意义采用单因素方差分析(ANOVA),p值小于0.05为差异显著。
讨 论
青蒿素是当前唯一尚未产生广泛耐药性的一线抗疟药物,也是为数不多的在整个疟原虫无性增殖IDC时期都有效的抗疟药物之一。目前迫切需要阐明ART的确切抗疟机制,以优化当前的ART用药方案并解决ART的耐药性,虽然已经有了暂时的缓解方案。由于疟原虫复杂的生命周期和复杂的药物作用机制(MOA),导致对ART抗疟机制的研究进展缓慢。目前,大多数研究集中在ART激活后对疟原虫蛋白的烷基化和共价相互作用的影响,而ART与疟原虫蛋白的非共价结合在抗疟过程中的作用以及生物学意义尚不明确,我们认为这种作用方式也不容忽视,有必要进行深入的研究。
在本研究中,利用活性光亲和探针在原位鉴定了恶性疟原虫IDC的环期、滋养体和裂殖体时期的ART蛋白靶标。我们研究了药物-蛋白结合方式,并进行了一系列的验证实验。结果表明,ART在疟原虫中主要被血红素激活后与蛋白质形成共价结合,但也存在非共价结合机制。一方面,这是由于疟原虫不同生命阶段抗疟作用途径的类型和重要性存在差异,以及与疟原虫生长阶段一致的靶标蛋白合成差异有关。另一方面,在结合方式(共价/非共价)上也有所不同,这可能是由于不同生长阶段的疟原虫细胞中含有不同水平的血红素,改变了ART的激活水平。本研究为全面研究ART共价和非共价结合靶标提供了重要的实验证据,这些靶标可能与ART的抗疟机制有关。此外,鉴于ART在疟原虫中的快速激活,尚不清楚有多少未激活形式的ART可逆结合,以及其对靶标蛋白功能的抑制程度,仍需要进一步深入研究。
然后,我们对所有靶标蛋白进行了生物信息学分析,并验证了可能参与ART抗疟作用的几个关键蛋白/生物学通路。我们的结果表明,ART可能通过抑制疟原虫蛋白合成,干扰糖酵解能量代谢,以及扰乱氧化还原相关的过程来发挥其抗疟作用。此外,我们还利用非靶向代谢组学技术研究了ART对疟原虫整体代谢过程的影响,但是仍需要更多的研究来确定ART最关键的抗疟靶标/生物学通路以及它们之间的关系。当前很多抗疟药物的MoAs仍需要进一步的阐明,本研究表明结合表型分析、靶点鉴定和代谢组学技术是阐明药物MoAS的一种有前景的方法。
结 论
本研究一方面为青蒿素靶标蛋白鉴定提供了丰富的数据,也为揭示青蒿素的抗疟机理提供了新的见解,这对于优化当前抗疟药物治疗方案和缓解青蒿素耐药性的产生具有重要的意义。
数据可用性
本文工作的数据已存入中国科学院生物信息中心/北京基因组研究所OMIX (https://ngdc.cncb.ac.cn/omix/preview/2DszKpI6)。使用的数据和脚本保存在GitHub (https://github.com/LabWang307/Antimalarial_Mechanism_ART)。所有补充材料(图表、表格、脚本、图形摘要、幻灯片、视频、中文翻译版本和更新材料)可以通过在线DOI或iMeta Science(http://www.imeta.science/)获取。
引文格式:
Gao, Peng, Jianyou Wang, Chong Qiu, Huimin Zhang, Chen Wang, Ying Zhang, Peng Sun, et al. 2024. Photoaffinity Probe‐Based Antimalarial Target Identification of Artemisinin in the Intraerythrocytic Developmental Cycle of Plasmodium falciparum. iMeta e176. https://doi.org/10.1002/imt2.176
作者简介

高鹏(第一作者)
● 中国中医科学院青蒿素研究中心助理研究员。
● 目前研究方向为青蒿素、氯喹等药物抗疟和耐药机制研究以及中药有效成分作用靶标鉴定,主持中国博士后基金,国家自然科学基金项目2项,相关学术成果发表于iMeta、Engineering、Military Medical Research、Drug Resistance Updates等国际一流期刊。

王建友(第一作者)
● 河南大学药学专业硕士。
● 主要研究课题为雷公藤红素等天然活性物质的潜在抗疟活性及其作用机制研究。相关学术成果已发表于Chinese Chemical Letters等期刊。

戴凌云(通讯作者)
● 南方科技大学临床副教授,深圳市人民医院特聘副研究员,广东省和深圳市科技创新高层次人才。
● 近年来一直从事功能蛋白靶标发现与药物靶向机制研究,基于MS-CETSA等多种功能蛋白组学技术解析肿瘤、衰老和感染等重大疾病发生发展进程中的关键靶标蛋白,系统性探究小分子药物、天然产物和代谢物的作用靶点及其多重药理作用。主要研究成果以第一或通讯作者身份发表于Cell、Cell Systems、Nature Protocols、Annual Review of Biochemistry、Acta Pharmaceutica Sinica B等国际一流期刊。

韩光(通讯作者)
● 河南大学药学院教授,博士生导师,抗病毒性传染病创新药物全国重点实验室(河南大学)常务副主任,河南省高层次人才,河南省“中药药效物质与创新药物研究”创新型科技团队带头人。
● 研究方向为天然活性成分研究与新药开发及基于系统生物学的纳米给药系统研究,主持国家自然科学基金、国家十一五重大新药创制子课题及重点科技攻关项目等20余项。研究成果发表于Cell Communication and Signaling、International journal of pharmaceutics等国际权威期刊。
更多推荐
(▼ 点击跳转)
高引文章 ▸▸▸▸
iMeta | 引用7000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
高引文章 ▸▸▸▸
iMeta | 德国国家肿瘤中心顾祖光发表复杂热图(ComplexHeatmap)可视化方法
高引文章▸▸▸▸
iMeta | 高颜值绘图网站imageGP+视频教程合集

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

2卷2期封底

2卷4期封底

3卷1期封底
期刊简介
“iMeta” 是由威立、肠菌分会和本领域数百位华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表原创研究、方法和综述以促进宏基因组学、微生物组和生物信息学发展。目标是发表前10%(IF > 15)的高影响力论文。期刊特色包括视频投稿、可重复分析、图片打磨、青年编委、前3年免出版费、50万用户的社交媒体宣传等。2022年2月正式创刊发行!
联系我们
iMeta主页:http://www.imeta.science
出版社:https://onlinelibrary.wiley.com/journal/2770596x
投稿:https://mc.manuscriptcentral.com/imeta
邮箱:office@imeta.science
